
I have always been fascinated by ruminants and specifically their rumens. During my undergrad years I had the chance to do an independent study into in vitro analysis of feeds and the rumen microbiota with the legendary Peter Van Soest (who by the way grew up on a dairy farm in Snohomish, WA and graduated from WSU with both his BS and MS in the 1950’s). One of the more fascinating aspects of the rumen milieu has to do with the protozoal and viral components. However, it has been somewhat difficult over the years to gain much insight into the roles these organisms play within the rumen.
My understanding of rumen protozoa has primarily been linked to their role as “canaries in the coal mine.” If the protozoa are missing or dead, the rumen microbiota must be suffering a collapse. In fact, this perspective was informed by Dr. Van Soest pointing out that valuation of the number and activity of protozoa in the ruminal fluid provides a sensitive indicator of the normality of the sample.1 This is easily accomplished by microscopic examination (×40 to ×100) of a drop of fresh, warm fluid without special stains. In very active fluid samples, the largest protozoa can be seen with the naked eye. They are detectable in a tube as small gray specks of material in active motion in the fluid, and they tend to localize above the sedimented particulate matter. Both ciliate and flagellate forms of varying sizes and shapes can be observed microscopically, with ciliates usually outnumbering the flagellates. The importance of the protozoa from a clinical viewpoint is their sensitivity to abnormalities in the fluid milieu.
Normal ruminal fluid should contain protozoa in a wide variety of sizes, in large numbers that are easy to see, and with active motility. Numbers of protozoa are reduced in inactive fluid samples. Larger species are more susceptible to abnormalities; thus, a predominance of small protozoa would suggest a mild indigestive disturbance. Although rumen protozoa can attenuate a sharp decrease of rumen pH in animals fed high-concentrate diets (likely attributable to their ability to uptake sugars and sequester them as glycogen), all protozoa are killed off when the ruminal pH drops below 5. Therefore, a recent bout of acidosis results in lack of protozoal activity, even if the pH has subsequently risen back into the normal range. Very recent disturbances of the fluid may result in the observation of a large number of dead protozoa.
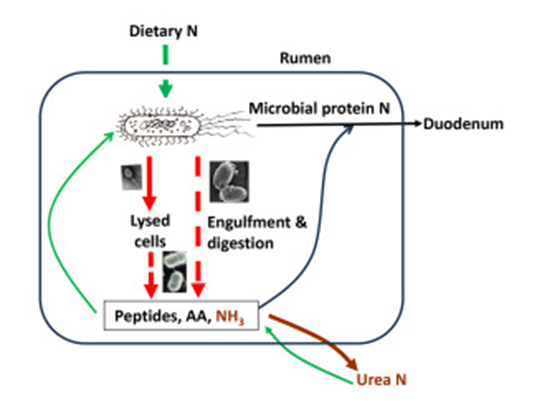
Even though it is established that the protozoa are normal inhabitants of a healthy ruminant’s ruminal fluid, their specific functions have historically been less understood when compared with their prokaryotic cohabitants and their presence does not appear to be a prerequisite of normal digestive activity.2 A recent paper in JDS Communications (Yu et al., 2024) provides a mini-review of our current understanding of rumen protozoa and viruses. The graphical abstract below and the paper’s summary highlight that protozoa and viruses (or phages) within the rumen act as predators, contributing to the intraruminal recycling of microbial protein. Specifically, protozoa engulf microbial cells and degrade their protein, releasing peptides, amino acids, and ammonia. While other microbes can use some of these products to resynthesize protein, considerable amounts are converted to urea and excreted. Viruses infect numerous rumen microbial cells of their hosts. They lyse their host cells through their lytic life cycle, making their cellular protein available for degradation by proteolytic bacteria. Elimination of rumen protozoa increases microbial protein supply to the small intestines by up to 30%, but it remains to be determined to what extent rumen viruses can potentially lower dietary nitrogen efficiency in dairy animals.
Due to their large size, protozoa constitute up to 50% of the rumen microbial biomass (Andersen et al., 2023).
Some rumen protozoa digest dietary fiber as evidenced by decreased fiber digestibility in defaunated animals.
Additionally, protozoa enhance bacterial fiber digestion, probably by scavenging the free oxygen entering the rumen and thus helping maintain anaerobiosis, a requirement for strictly anaerobic fibrolytic bacteria. Through fermentation, rumen protozoa produce volatile fatty acids (VFA), contributing to the major metabolizable energy (ME) for ruminants. Ultimately, the substantial protozoan biomass and high digestibility of rumen protozoa underscore their importance as a microbial protein source for the host. However, as mentioned above their substantive contribution to the intraruminal recycling of microbial protein concurrently lowers microbial protein supply to the host and reduces dietary N utilization efficiency.
Dairy cows emit CH4 during rumen fermentation and metaanalyses have revealed a linear relationship between CH4 emissions and protozoa abundance, and demonstrated that eliminating rumen protozoa could decrease CH4 emissions by 11%, while decreasing organic matter digestibility by 7% (Guyader et al., 2014; Newbold et al., 2015). The stimulation of CH4 by rumen protozoa stems from their ability to produce hydrogen during fermentation within their hydrogenosomes or in their cytosol, and to form ecto- and endo-symbiotic relationships with methanogens. Importantly, dietary interventions that reduce rumen protozoa can mitigate CH4 emissions (Almeida et al., 2021).
Viruses, including those infecting bacteria, archaea, fungi, and protozoa are abundant in the rumen, reaching 107 to 1010 virions/mL of rumen fluid (Lobo and Faciola, 2021). These viruses have 2 distinct lifecycles: the lytic and the lysogenic cycles. Lytic viruses lyse their host cells to release progeny viruses for subsequent rounds of infection. The viral lysis also releases host cellular components, increasing nutrient cycling in the rumen. In the lysogenic cycle, viruses insert their genomes into the host genomes as prophages, leading to lysogeny. Lysogenic viruses can provide
their host with new metabolic capabilities, buttressing ecological fitness and potentially facilitating host evolution. Therefore, rumen viruses can profoundly affect the rumen microbiome and its function, with the potential for the rumen virome to affect the diversity, composition, metabolism, and functions of the rumen ecosystem.
Overall, rumen protozoa and viruses, acting as predators within the rumen microbiome, remain less understood compared with bacteria and methanogens, primarily due to challenges in their definitive characterization through culturing. Recent omics studies have greatly advanced our understanding of these rumen microbes, providing new insights into their crucial roles in shaping the rumen microbiome and its functions. Integrating genomics, transcriptomics, and proteomics holds promise for identifying specific targets to control rumen protozoa effectively without collateral damage. In the context of dairy production, exploring certain rumen viruses may offer novel strategies to control undesirable rumen microbes, including methanogens and protozoa. Specifically, the phages infecting rumen protozoa, particularly those highly bacterivorous and those infecting hyper ammonia-producing bacteria, hold the potential to reduce intraruminal recycling of microbial protein and deamination, thereby enhancing dietary N efficiency.
- 1van Soest PJ. Nutritional ecology of the ruminant. Ithaca: Comstock Publishing Associates, 1994.
- 2Williams AG, Coleman GS. Role of protozoa in the rumen In: Williams AG, Coleman GS, eds. The rumen protozoa. New York: Springer-Verlag, 1992;317-374.